
Articles
- Page Path
- HOME > Osong Public Health Res Perspect > Volume 4(1); 2013 > Article
-
Original Article
A Comparison of Subtyping Methods for DifferentiatingSalmonella enterica Serovar Enteritidis Isolates Obtained from Food and Human Sources - Ji-Yeon Hyeona,b, Jung-Whan Chona, Jun-Ho Parka, Moo-Sang Kima, Young-Hee Oha, In-Soo Choia, Kun-Ho Seoa
-
Osong Public Health and Research Perspectives 2013;4(1):27-33.
DOI: https://doi.org/10.1016/j.phrp.2012.12.005
Published online: February 28, 2013
aCollege of Veterinary Medicine, Konkuk University, Seoul, Korea.
bDivision of Vaccine Research, Korea National Institute of Health, Osong, Korea.
- *Corresponding author. E-mail: bracstu3@konkuk.ac.kr
• Received: November 6, 2012 • Revised: December 15, 2012 • Accepted: December 19, 2012
Copyright ©2013, Korea Centers for Disease Control and Prevention
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
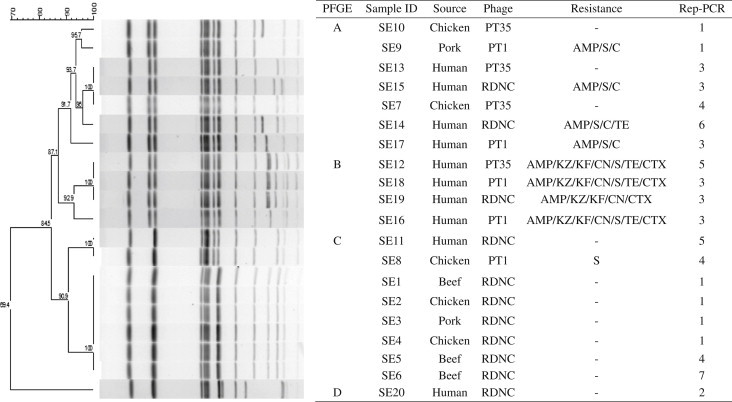
Figure & Data
References
Citations
Citations to this article as recorded by 
- Genomic and phylogenetic analysis of Salmonella enterica serovar Enteritidis strains linked to multiple outbreaks in Brazil
Guojie Cao, Maria Balkey, Qing Jin, Eric Brown, Marc Allard, Adma Nadja Ferreira de Melo, Geany Targino de Souza Pedrosa, Tereza Cristina Rocha Moreira de Oliveira, Marciane Magnani, Dumitru Macarisin
Letters in Applied Microbiology.2023;[Epub] CrossRef - Phenotypic and Genotypic Characterization of Salmonella Isolates Recovered from Foods Linked to Human Salmonellosis Outbreaks in Minas Gerais State, Brazil
Leandro L. Faula, Monica M. O.P. Cerqueira, Jovita E. G.C. Madeira, Marcelo R. Souza, Fernando N. Souza, Gláucia C.S. Amancio
Journal of Food Protection.2022; 85(1): 142. CrossRef - Comparison of conventional molecular and whole-genome sequencing methods for subtyping Salmonella enterica serovar Enteritidis strains from Tunisia
Boutheina Ksibi, Sonia Ktari, Houcemeddine Othman, Kais Ghedira, Sonda Maalej, Basma Mnif, Mohamed salah Abbassi, Laetitia Fabre, Faouzia Rhimi, Simon Le Hello, Adnene Hammami
European Journal of Clinical Microbiology & Infect.2021; 40(3): 597. CrossRef - Molecular characterisation of antimicrobial resistance determinants and class 1 integrons of Salmonella enterica subsp. enterica serotype Enteritidis strains from retail food in China
Yu Li, Xiaojuan Yang, Jumei Zhang, Shiyuan Yang, Shuhong Zhang, Moutong Chen, Liang Xue, Yu Ding, Haiyan Zeng, Qihui Gu, Youxiong Zhang, Xianhu Wei, Juan Wang, Qingping Wu
Food Control.2021; 128: 108191. CrossRef - Molecular typing of cephalosporin resistant serovars of Salmonella enterica from poultry and farm animals
A. Rafati Zomorodi, M. Rad, Gh. R. Hashemitabar, H. Salimizand
BULGARIAN JOURNAL OF VETERINARY MEDICINE.2020; 23(2): 178. CrossRef -
Molecular characterization of
Salmonella
isolates recovered from slaughtered poultry in Trinidad
Nitu Kumar, Krishna Mohan, Karla Georges, Francis Dziva, Abiodun A. Adesiyun
Journal of Food Processing and Preservation.2020;[Epub] CrossRef - Effectiveness of BOX-PCR in Differentiating Genetic Relatedness amongSalmonella entericaSerotype 4,[5],12:i:- Isolates from Hospitalized Patients and Minced Pork Samples in Northern Thailand
Kritchai Poonchareon, Chaiwat Pulsrikarn, Narong Nuanmuang, Phichaya Khamai
International Journal of Microbiology.2019; 2019: 1. CrossRef - A common Salmonella Enteritidis sequence type from poultry and human gastroenteritis in Ibagué, Colombia
Luz Clemencia Fandiño, Noel Verjan
Biomédica.2019; 39: 50. CrossRef - Distribution and Molecular Characterization of Campylobacter Species at Different Processing Stages in Two Poultry Processing Plants
Soo-Kyoung Lee, Hyun-Jung Park, Jin-Hee Lee, Jong-Soo Lim, Kun-Ho Seo, Eun-Jeong Heo, Young-Jo Kim, Sung-Hwan Wee, Jin-San Moon
Foodborne Pathogens and Disease.2017; 14(3): 141. CrossRef - Molecular Characterization of Salmonella from Beef Carcasses and Fecal Samples from an Integrated Feedlot and Abattoir in Mexico
Diana Ayala, Kendra Nightingale, Claudia Narvaez-Bravo, Mindy M. Brashears
Journal of Food Protection.2017; 80(12): 1964. CrossRef - Repetitive sequence-based PCR fingerprinting and the relationship of antimicrobial-resistance characteristics and corresponding genes among Salmonella strains from pig production
Teerarat Prasertsee, Nattakarn Khantaprab, Panuwat Yamsakul, Pannita Santiyanont, Nipa Chokesajjawatee, Prapas Patchanee
Asian Pacific Journal of Tropical Disease.2016; 6(5): 390. CrossRef - Prevalence, Seasonal Occurrence, and Antimicrobial Resistance of Salmonella spp. Isolates Recovered from Chicken Carcasses Sampled at Major Poultry Processing Plants of South Korea
Soo-Kyoung Lee, Dasom Choi, Hong-Seok Kim, Dong-Hyeon Kim, Kun-Ho Seo
Foodborne Pathogens and Disease.2016; 13(10): 544. CrossRef - Resistance of Strains Producing Extended-Spectrum β-Lactamases AmongSalmonellafrom Duck Carcasses at Slaughterhouses in Three Major Provinces of South Korea
Soo Kyoung Lee, Dasom Choi, Jung Whan Chon, Kun Ho Seo
Foodborne Pathogens and Disease.2016; 13(3): 135. CrossRef - High Occurrence of Extended-Spectrum β-Lactamase-ProducingSalmonellain Broiler Carcasses from Poultry Slaughterhouses in South Korea
Jung-Whan Chon, Hae-In Jung, Min Kuk, Young-Ji Kim, Kun-Ho Seo, Soo-Ki Kim
Foodborne Pathogens and Disease.2015; 12(3): 190. CrossRef - Comparison of four molecular methods to typeSalmonellaEnteritidis strains
Fábio Campioni, André Pitondo-Silva, Alzira M.M. Bergamini, Juliana P. Falcão
APMIS.2015; 123(5): 422. CrossRef - Modified Method of Multilocus Sequence Typing (MLST) for Serotyping inSalmonellaSpecies
Young-Sun Yun, Su-Jin Chae, Hye-Young Na, Gyung Tae Chung, Cheon-Kwon Yoo, Deog-Yong Lee
Journal of Bacteriology and Virology.2015; 45(4): 314. CrossRef - Molecular Characterization of Salmonella Paratyphi B dT + and Salmonella Heidelberg from Poultry and Retail Chicken Meat in Colombia by Pulsed-Field Gel Electrophoresis
Pilar Donado-Godoy, Barbara A. Byrne, Michael Hume, Maribel León, Enrique Pérez-Gutiérrez, Martha J. Vives Flores, Viviana Clavijo, Ángela Holguin, Juan J. Romero-Zuňiga, Ricardo Castellanos, Mcallister Tafur, Woutrina A. Smith
Journal of Food Protection.2015; 78(4): 802. CrossRef - Prevalence Analysis and Molecular Characterization of Salmonella at Different Processing Steps in Broiler Slaughter Plants in South Korea
Hyun‐Jung Park, Jung‐Whan Chon, Jong‐Soo Lim, Kun‐Ho Seo, Young‐Jo Kim, Eun‐Jeong Heo, Sung‐Hwan Wee, Kidon Sung, Jin‐San Moon
Journal of Food Science.2015;[Epub] CrossRef - Characterization of Escherichia coli–Producing Extended-Spectrum β-Lactamase (ESBL) Isolated from Chicken Slaughterhouses in South Korea
Jong-Soo Lim, Da-Som Choi, Young-Jo Kim, Jung-Whan Chon, Hong-Seok Kim, Hyun-Jung Park, Jin-San Moon, Sung-Hwan Wee, Kun-Ho Seo
Foodborne Pathogens and Disease.2015; 12(9): 741. CrossRef - Comparative Analysis of Subtyping Methods against a Whole-Genome-Sequencing Standard for Salmonella enterica Serotype Enteritidis
Xiangyu Deng, Nikki Shariat, Elizabeth M. Driebe, Chandler C. Roe, Beth Tolar, Eija Trees, Paul Keim, Wei Zhang, Edward G. Dudley, Patricia I. Fields, David M. Engelthaler, N. A. Ledeboer
Journal of Clinical Microbiology.2015; 53(1): 212. CrossRef - Prevalence, characterization, and antimicrobial susceptibility of Salmonella Gallinarum isolated from eggs produced in conventional or organic farms in South Korea
Soo-Kyoung Lee, Jung-Whan Chon, Kwang-Young Song, Ji-Yeon Hyeon, Jin-San Moon, Kun-Ho Seo
Poultry Science.2013; 92(10): 2789. CrossRef
 PubReader
PubReader Cite
Cite
