
Articles
- Page Path
- HOME > Osong Public Health Res Perspect > Volume 2(1); 2011 > Article
-
Original Article
Genotypic Characterization ofVibrio vulnificus Clinical Isolates in Korea - Hye Sook Jeonga, Jun Young Kimb, Se Mi Jeonb, Mi Sun Parkb, Seong Han Kimb
-
Osong Public Health and Research Perspectives 2011;2(1):8-14.
DOI: https://doi.org/10.1016/j.phrp.2011.04.008
Published online: April 27, 2011
aDivision of Enteric and Hepatitis Virus, Center for Infectious Diseases, Korea National Institute of Health, Osong, Korea
bDivision of Enteric Bacterial Infections, Center for Infectious Diseases, Korea National Institute of Health, Osong, Korea
- ∗Corresponding author. kking@korea.kr
- 1Hye Sook Jeong and Jun Young Kim contributed equally to this study.
• Received: February 11, 2011 • Revised: March 29, 2011 • Accepted: April 1, 2011
© 2011 Published by Elsevier B.V. on behalf of Korea Centers for Disease Control and Prevention.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
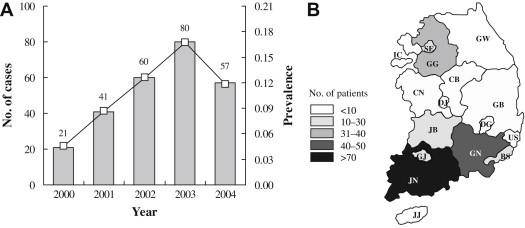



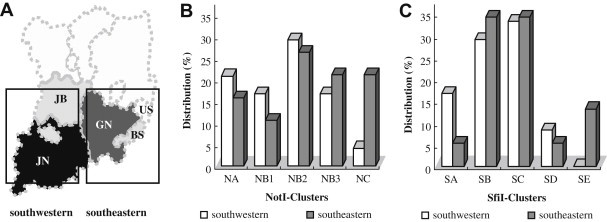
Figure & Data
References
Citations
Citations to this article as recorded by 
- Effect of Seawater Temperature Increase on the Occurrence of Coastal Vibrio vulnificus Cases: Korean National Surveillance Data from 2003 to 2016
Jungsook Kim, Byung Chul Chun
International Journal of Environmental Research an.2021; 18(9): 4439. CrossRef - PCR-based evidence showing the presence of Vibrio vulnificus in wound infection cases in Mangaluru, India
Caroline D’Souza, Ballamoole Krishna Kumar, Sachidananda Kapinakadu, Ranjith Shetty, Indrani Karunasagar, Iddya Karunasagar
International Journal of Infectious Diseases.2018; 68: 74. CrossRef - Genotypic Diversity and Population Structure of Vibrio vulnificus Strains Isolated in Taiwan and Korea as Determined by Multilocus Sequence Typing
Hye-Jin Kim, Jae-Chang Cho, Paul J Planet
PLOS ONE.2015; 10(11): e0142657. CrossRef - The Road Less Traveled
Chaeshin Chu
Osong Public Health and Research Perspectives.2011; 2(1): 1. CrossRef
 PubReader
PubReader Cite
Cite




