
Articles
- Page Path
- HOME > Osong Public Health Res Perspect > Volume 5(1); 2014 > Article
-
Original Article
Role of Active and Inactive Cytotoxic Immune Response in Human Immunodeficiency Virus Dynamics - Hernan Dario Toro Zapataa, Angelica Graciela Caicedo Cassob, Derdei Bicharac, Sunmi Leed
-
Osong Public Health and Research Perspectives 2014;5(1):3-8.
DOI: https://doi.org/10.1016/j.phrp.2014.01.001
Published online: January 31, 2014
aLicenciatura en Matemáticas, Universidad del Quindío, Quindío, Colombia
bDepartamento de Matematicas, Universidad del Valle, Cali, Colombia
cMathematical, Computational and Modeling Sciences Center, Arizona State University, Tempe, AZ, USA
dDepartment of Applied Mathematics, Kyung Hee University, Yongin, Korea
- ∗Corresponding author. sunmilee@khu.ac.kr
• Received: October 12, 2013 • Revised: January 10, 2014 • Accepted: January 13, 2014
© 2014 Published by Elsevier B.V. on behalf of Korea Centers for Disease Control and Prevention.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
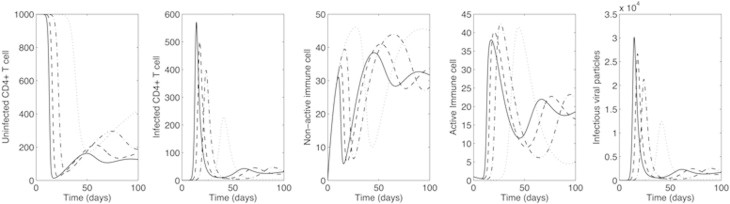

Figure & Data
References
Citations
Citations to this article as recorded by 
- A Multi-Scale Model for the Spread of HIV in a Population Considering the Immune Status of People
Sol de Amor Vásquez-Quintero, Hernán Darío Toro-Zapata, Dennis Alexánder Prieto-Medellín
Processes.2021; 9(11): 1924. CrossRef - Qualitative investigation of cytolytic and noncytolytic immune response in an HBV model
John G. Alford, Stephen A. McCoy
Involve, a Journal of Mathematics.2020; 13(3): 455. CrossRef - Mathematical Model Describing HIV Infection with Time-Delayed CD4 T-Cell Activation
Hernán Darío Toro-Zapata, Carlos Andrés Trujillo-Salazar, Edwin Mauricio Carranza-Mayorga
Processes.2020; 8(7): 782. CrossRef - The Role of Immune Response in Optimal HIV Treatment Interventions
Hernán Toro-Zapata, Angélica Caicedo-Casso, Sunmi Lee
Processes.2018; 6(8): 102. CrossRef - Evaluación teórica de estrategias óptimas y sub-óptimas de terapia antirretroviral para el control de la infección por VIH
Hernán Darío Toro-Zapata, Carlos Andrés Trujillo-Salazar, Dennis Alexánder Prieto-Medellín
Revista de Salud Pública.2018; 20(1): 117. CrossRef - Summing Up Again
Hae-Wol Cho, Chaeshin Chu
Osong Public Health and Research Perspectives.2014; 5(4): 177. CrossRef - Roll the Dice
Hae-Wol Cho, Chaeshin Chu
Osong Public Health and Research Perspectives.2014; 5(5): 243. CrossRef - Journal Publishing: Never Ending Saga
Hae-Wol Cho, Chaeshin Chu
Osong Public Health and Research Perspectives.2014; 5(1): 1. CrossRef
 PubReader
PubReader Cite
Cite

