
Articles
- Page Path
- HOME > Osong Public Health Res Perspect > Volume 4(1); 2013 > Article
-
Original Article
A Comparison of Subtyping Methods for DifferentiatingSalmonella enterica Serovar Enteritidis Isolates Obtained from Food and Human Sources - Ji-Yeon Hyeona,b, Jung-Whan Chona, Jun-Ho Parka, Moo-Sang Kima, Young-Hee Oha, In-Soo Choia, Kun-Ho Seoa
-
Osong Public Health and Research Perspectives 2013;4(1):27-33.
DOI: https://doi.org/10.1016/j.phrp.2012.12.005
Published online: February 28, 2013
aCollege of Veterinary Medicine, Konkuk University, Seoul, Korea.
bDivision of Vaccine Research, Korea National Institute of Health, Osong, Korea.
- *Corresponding author. E-mail: bracstu3@konkuk.ac.kr
• Received: November 6, 2012 • Revised: December 15, 2012 • Accepted: December 19, 2012
Copyright ©2013, Korea Centers for Disease Control and Prevention
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Abstract
-
Purpose:
- To evaluate the abilities of these subtyping methods, we distinguished Salmonella Enteritidis (S. Enteritidis) isolated from food products and human clinical samples between 2009 and 2010 in Seoul using five subtyping methods.
-
Methods:
- We determined the subtypes of 20 S. Enteritidis isolates from food and human sources using phage typing, antimicrobial susceptibility, pulsed-field gel electrophoresis (PFGE), repetitive sequence-based PCR (rep-PCR), and multi-locus sequence typing (MLST).
-
Results:
- A total of 20 tested isolates were differentiated into six antimicrobial susceptibility patterns, three different phage types, four different PFGE profiles, seven rep-PCR patterns, and one MLST type. Food isolates were considerably more susceptible to antibiotics than human isolates. We were best able to discriminate among S. Enteritidis isolates using rep-PCR, and obtained the highest Simpson’s diversity index of 0.82, whereas other methods produced indices that were less than 0.71. PFGE pattern appeared to be more related to antimicrobial resistance and phage types of S. Enteritidis isolates than rep-PCR. MLST revealed identical alleles in all isolates at all seven loci examined, indicating no resolution.
-
Conclusion:
- The results of this study suggest that rep-PCR provided the best discriminatory power for phenotypically similar S. Enteritidis isolates of food and human origins, whereas the discriminatory ability of MLST may be problematic because of the high sequence conservation of the targeted genes.
- Nontyphoidal Salmonella enterica is a major cause of foodborne illness worldwide [1,2]. Most human cases of nontyphoidal Salmonella result from the consumption of contaminated foods of animal origin, especially poultry meat and eggs [3–5]. The S. enterica serotypes Typhimurium, Enteritidis, and Newport were the most common serotypes identified among human infection pathogens commonly transmitted through food, according to preliminary FoodNet data for 2005 [2].
- Salmonella typing technologies are essential for bacterial source tracking and to determine the distribution of pathogens that have been isolated from infected people [6]. Traditional typing methods based on phenotypic traits, such as biotyping, antibiotic susceptibility profiles, serotyping, and phage typing, provide insufficient information for epidemiological purposes [6,7]. Molecular subtyping methods have revolutionized the identification of microbial strains, but most of them have not been internationally standardized [7]. Subtyping methods have been developed based on three main mechanisms of discrimination: (1) restriction analysis of bacterial DNA, (2) polymerase chain reaction (PCR) amplification of particular genetic targets, and (3) the identification of DNA sequence polymorphism at specific loci in the genome [6].
- Pulsed-field gel electrophoresis (PFGE) is a form of restriction fragment length polymorphism analysis typing, in which restriction patterns of whole bacterial genomes are analyzed and compared [6,7]. PFGE is used by the PulseNet program, a molecular subtyping network for foodborne bacterial disease surveillance at the U.S. Centers for Disease Control and Prevention (CDC), to identify widespread outbreaks of bacterial foodborne illness [8,9].
- Repetitive sequence-based PCR (rep-PCR) is an amplification-based method that utilizes the repeated DNA sequence elements distributed throughout the genomes of many bacterial species [6]. Rep-PCR uses noncoding repetitive sequence primers to produce copies of DNA fragments [10]. Differences in the resulting banding patterns due to differences in the number and size of amplified repetitive elements and can be compared to determine the genetic relatedness of microbial strains [6,10].
- In recent years, powerful sequencing facilities and the availability of genome sequences have allowed the development of new typing methods such as multilocus sequence typing (MLST) [11]. In MLST, the variability in a relatively small part of the genome due to mutation or recombination events is investigated through the comparison of nucleotide base changes in multiple genes with conserved sequences, such as housekeeping genes [8,12–14]. MLST is a relatively expensive method that may not be available for use in many clinical laboratories or for routine surveillance [12]. However, it has been developed for a number of clinically important bacterial pathogens, including Salmonella spp., and can be useful as a discriminatory typing method for Salmonella spp. [13].
- There are many subtyping methods that have been developed as described above, but only PFGE is often considered the “gold standard” for molecular typing methods of bacterial foodborne pathogens [6,8,15]. In addition, few studies that evaluated the ability of sub-typing methods based on different mechanisms of discrimination to differentiate phenotypically similar but epidemiologically unrelated isolates were reported. Therefore, we determined the subtypes S. Enteritidis isolates from food and human sources using two phenotypic subtyping methods (phage typing and antimicrobial susceptibility) and three genotypic subtyping methods (PFGE, rep-PCR, and MLST) and compared their ability to distinguish among S. Enteritidis isolates.
Introduction
- 2.1. Bacterial isolates
- We used a total of 20 S. Enteritidis isolates isolated from human fecal samples (n = 10) and retail meats (n = 10). The 10 retail-meat isolates were from beef (n = 3), pork (n = 2), and chicken (n = 5) that were obtained from retail markets in Seoul, South Korea, in 2009 and 2010. Ten human isolates were randomly selected from the culture collection of the Seoul Research Institute of Public Health and Environment (SIHE; Gwachon, South Korea). These samples were isolated from sporadic diarrheal patients in Seoul, South Korea, between 2009 and 2010. SIHE confirmed that the isolates were Salmonella by using VITEK Gram-negative identification cards (bioMérieux, Durham, NC) and serotyped using commercial Difco antisera (BD, Sparks, MD, USA).
- 2.2. Phage typing
- All 20 S. Enteritidis isolates were phage typed using the standardized CDC method [16,17]. Phage typing was performed at the Animal, Plant, and Fisheries Quarantine and Inspection Agency (Anyang, South Korea).
- 2.3. Antibiotic susceptibility test
- The antibiotic susceptibilities of S. Enteritidis isolates were determined with the disk diffusion method, as recommended by the Clinical and Laboratory Standards Institute [18]. Sensi-Disc Antimicrobial Susceptibility Test Discs (Oxoid, Basingstoke, UK) were used with the following antibiotics: 10 μg ampicillin, 30 μg amikacin, 30 μg chloramphenicol, 30 μg cephalothin, 5 μg ciprofloxacin, 10 μg gentamicin, 10 μg streptomycin, 25 μg sulfamethoxazole/trimethoprim, 30 μg tetracycline, 30 μg cefazolin, 30 μg amoxicillin/clavulanic acid, 30 μg cefepime, 30 μg cefoxitin, 30 μg cefotaxim, 5 μg enrofloxacin, 10 μg norfloxacin, and 10 μg imipenem. The diameter of inhibition zones was recorded, and samples were scored as sensitive, intermediate, and resistant according to the scoring system recommended by the Clinical and Laboratory Standards Institute.
- 2.4. PFGE
- DNA and agarose were prepared for PFGE as described in previous studies [19,20]. For restriction endonuclease digestion, two 1-mm-thick slices of each plug were incubated at 37° C for 1.5 h with 30 U of NotI enzyme (Takara Bio Inc., Otsu, Shiga, Japan) in 100 mL of the appropriate restriction enzyme buffer. The restriction fragments were separated by electrophoresis in 0.5M Tris borate–EDTA buffer at 14 °C for 18 h using a Chef Mapper electrophoresis system (Bio-Rad, Hercules, CA, USA) with pulse times between 2.16 and 54.17 seconds. The gels were stained with ethidium bromide, and DNA bands were visualized with UV transillumination (Bio-Rad). Salmonella serovar Braenderup ATCC BAA 664 was used as the control strain and digested with 30 U of XbaI (Takara Bio Inc.). Interpretation of DNA fingerprint patterns was accomplished using Bionumerics 4.0 software (Applied Maths, Austin, TX, USA). The banding patterns were compared using Dice coefficients with a 1.5% band position tolerance. Patterns with no noticeable differences were considered indistinguishable and were assigned the same PFGE pattern designation.
- 2.5. Rep-PCR using DiversiLab
- S. Enteritidis was cultured on nutrient agar (Difco) for 24 h at 37° C. DNA from each isolate was extracted using UltraClean Microbial DNA Isolation Kits (MoBio Laboratories, Solana Beach, CA, USA) based on the manufacturer’s instructions. Genomic DNA samples were quantified using a NanoDrop 2000 UV spectrophotometer (Thermo Scientific, Wilmington, DE, USA) at 260 nm. For PCR reactions, 2 μL of genomic DNA (approximately 25 ng/μL) was amplified using the DiversiLab Salmonella Kit (bioMérieux Inc.). The final mixture (23 μL) contained 0.5 μL (or 2.5 U) of AmpliTaq polymerase (Applied Biosystems, Foster City, CA, USA), 2.5 μL of 10× GeneAMP PCR Buffer I (Applied Biosystems), 2 μL kit-supplied primer mix, and 18 μL of the kit-supplied rep-PCR master mix (MM1). Thermal cycling parameters were as follows: initial denaturation at 94 °C for 2 min; followed by 35 cycles of denaturation at 94 °C for 30 seconds, annealing at 50 °C for 30 seconds, extension at 70 °C for 90 seconds; and a final extension at 70 °C for 3 minutes. The rep-PCR products were separated and detected by a micro-fluidics chip and Agilent model 2100 Bioanalyzer (Agilent Technologies Inc., Palo Alto, CA, USA). DNA fingerprint patterns were generated as electropherogram, which were automatically exported to the DiversiLab software for analysis. Both band position and intensity of all samples were analyzed and compared with web-based DiversiLab software (version 3.3) using the extended Jaccard coefficient. Distance matrices and the unweighted pair-group method with arithmetic mean were used to create a dendrogram [21]. The isolates were categorized as follows: indistinguishable isolates had >97% similarity with no banding differences and no variation in intensities of individual bands; similar isolates had 95–97% similarity and one or two different bands difference; different isolates had <95% similarity and two or more different bands [21]. Isolates were assigned unique rep-PCR types unless classed as indistinguishable or similar using the above criteria.
- 2.6. MLST
- MLST was performed according to the method described in previous studies [12,13,22–30] (http://mlst.ucc.ie/mlst/dbs/Senterica). DNA was extracted using an UltraClean Microbial DNA Isolation Kit. PCR reactions were performed in 20 μL reaction mixtures in a thermocycler (Biometra, Göttingen, Germany). For PCR reactions, 0.5 μM DNA template was added to 50 μL PCR mixture consisting of Pyrobest DNA polymerase (5 U/μL, 0.5 μL), 10× Pyrobest buffer II (Takara Bio Inc.), 1 μM of each primer (http://mlst.ucc.ie/mlst/dbs/Senterica), and dNTP mixture (2.5 mM each). The mixture was initially denatured at 98 °C for 5 minutes followed by 40 cycles of 98 °C for 10 seconds, 55 °C for 30 seconds, 72 °C for 1 minute, and finally elongated at 72 °C for 5 minutes. The PCR products were separated on 2% agarose gels. TIFF images of agarose gels were generated using the GelDoc XR+ gel documentation system (Bio-Rad) and Image Lab 3.0 Software (Bio-Rad). After PCR product purification, the DNA sequences of clones were analyzed by the Custom Oligonucleotide Synthesis Manufacture Office (Seoul, South Korea). Allele numbers were assigned an MLST type after the distinct allele sequences were submitted via the Internet to the dedicated database (http://mlst.ucc.ie/mlst/dbs/Senterica).
- 2.7. Data analysis
- The diversity of PFGE, rep-PCR, and MLST was assessed using Simpson’s index (D). Confidence intervals were calculated as described in a previous study [31]. We calculated Simpson’s index as: where N is the total number of isolates in the sample population, s is the total number of types, and nj is the number of isolates belonging to the jth type.
Materials and Methods
- 3.1. Antibiotic susceptibility profiles
- Six antibiotic susceptibility patterns of S. Enteritidis isolates are shown in Figure 1, and the Simpson’s index of this method was 0.68 (Table 1). Twenty percent of food isolates and 70% of human isolates were resistant to at least one of the 16 antibiotics tested. The highest resistance rate was observed for ampicillin and streptomycin (40%), followed by chloramphenicol, cephalothin, gentamicin, tetracycline, cefazolin, and cefotaxim (20%). Eight of the 20 isolates (40%) were resistant to two or more antibiotics, and seven of these were human isolates (Figure 1). Among the eight isolates resistant to two or more antibiotics, SE12, SE16, SE18, and SE 19 were resistant to ampicillin–cefazolin–cephalothin–gentamicin–cefotaxim–(streptomycin–tetracycline), and the other four were resistant to ampicillin–streptomycin–chloramphenicol (Figure 1).
- 3.2. Phage type
- The S. Enteritidis isolate phage types are shown in Figure 1. The Simpson’s index from the results of this method was the lowest, at 0.62 (Table 1). We found a total of three phage types, PT1, PT35, and reaction does not confirm (RDNC), among the 20 isolates tested. These phage types were as follows: RDNC (55%, six food and five human isolates), PT35 (25%, two food and three human isolates), and PT1 (20%, two food and two human isolates).
- 3.3. PFGE and rep-PCR patterns
- A total of four types (A through D) of PFGE pattern were generated from the 20 S. Enteritidis isolates (Figure 1), and the Simpson’s index was 0.71 (Table 1). In rep-PCR pattern, a total of seven types (1 through 7) were generated (Figure 1), and the Simpson’s index was 0.82 (Table 1). Among the PFGE patterns, PFGE type C was the largest group with seven food isolates and one human isolate (Figure 1). Seven of eight isolates of PFGE type C with 100% similarity were phage type RDNC and not resistant to antibiotics, and they were included in rep-PCR types 1, 4, and 7. All four human isolates of PFGE type B had a similar antimicrobial resistance pattern, ampicillin–cefazolin–cephalothin–gentamicin–cefotaxim, and three of them showed 100% similarity in PFGE and separated to rep-PCR types 3 and 5 (Figure 1). PFGE type A included four human and three food isolates that were either not resistant to antibiotics or resistant to ampicillin–streptomycin–chloramphenicol (Figure 1). PFGE type A isolates were differentiated into rep-PCR types 1, 3, 4, and 6 (Figure 1).
- Among rep-PCR patterns, rep-PCR types 1 and 3 were large groups, and four isolates of rep-PCR type 1 were phage type RDNC and not resistant to antibiotics and were included in PFGE type C. In addition, three isolates of rep-PCR type 3 were resistant to ampicillin–cefazolin–cephalothin–gentamicin–cefotaxim and were included in PFGE type B. However, rep-PCR types 4 and 5 did not show a similar phenotypic pattern, and rep-PCR types 2, 6, and 7 had one isolate.
- 3.4. MLST analysis
- Seven housekeeping genes (aroC, dnaN, hemD, hisD, purE, sucA, and thrA) were compared using MLST analysis (Table 1). All isolates were assigned to MLST 11 in the dedicated database (http://mlst.ucc.ie/mlst/dbs/Senterica), resulting in the Simpson’s index (D) of zero (Table 1). All isolates possessed identical alleles at all seven loci; aroC allele type 5, dnaN allele type 2, hemD allele type 3, hisD allele type 7, purE allele type 6, sucA allele type 6, and thrA allele type 11.
Results
- Given the public health hazard posed by S. Enteritidis, it is important to be able to quickly describe outbreaks, trace transmission routes, and define relationships between human and food isolates [32,33].We characterized S. Enteritidis isolates (n = 20) from food and human sources by antibiotic susceptibility, phage typing, PFGE, rep-PCR, and MLST. We then evaluated the correlations between the types formed using the results of each subtyping method. Six antimicrobial susceptibility patterns, three phage types, four PFGE types, and seven rep-PCR types were identified among the 20 isolates. MLST typing had no discriminatory power, as only one MLST type was observed.
- We also investigated the typing methods as tools for determining the source, either human or food, of S. Enteritidis isolates. Antibiotic resistance to two or more antibiotics was more common in human isolates than in food isolates. PFGE and rep-PCR tended to generate human or animal-specific clustering. Many of the human isolates possessed similar PFGE patterns (type A and B), and food and human isolates were not included in same type in rep-PCR pattern.
- The discriminatory power of five subtyping methods was measured by calculation of Simpson’s index. Simpson’s index is commonly used as an estimate of the discriminatory ability of subtyping methods, and it is a measure of the probability that two epidemiologically unrelated isolates will be characterized as being “different” by the typing method under evaluation [32,34]. The highest Simpson’s index was obtained from rep-PCR followed by PFGE, antibiotic susceptibility, phage typing, and MLST. In a previous study, the semiautomated rep-PCR (DiversiLab) patterns have a limited ability to discriminate some serotypes of Salmonella from different sources [35,36]. In addition, PFGE and rep-PCR exhibited a similar discriminatory ability in patient and food samples from a large food-borne outbreak of S. Enteritidis [33]. However, in the current study using epidemiologically unrelated isolates, rep-PCR showed the best discriminatory power with the highest Simpson’s index.
- PFGE pattern appeared to be more related to antimicrobial resistance profiles and phage types of S. Enteritidis isolates than rep-PCR, and Simpson’s index was higher in rep-PCR than in PFGE (Figure 1 and Table 1). S. Enteritidis isolates with similar antibiotic resistance pattern were indistinguishable in PFGE types A, B, and C. However, these isolates were separated into rep-PCR types 1, 3, and 6 (PFGE type A), types 3 and 5 (PFGE type B), and types 1, 4, 5, and 7 (PFGE type C). These results of PFGE pattern in this study are in agreement with those of previous studies by Foley et al [8] and Harbottle et al [13]. S. Typhimurium isolates with indistinguishable PFGE patterns exhibited resistance to at least five antimicrobials [8], and S. Newport isolates with indistinguishable PFGE patterns exhibited similar antibiotic resistance patterns [13]. These results indicate that S. Enteritidis isolates with similar antimicrobial susceptibility patterns might be indistinguishable by PFGE. This limitation can be overcome by the use of a second enzyme for PFGE analysis or a combination of rep-PCR, further increasing the ability of researchers to differentiate between S. Enteritidis isolates that have similar phenotypes [8,37].
- In this study, all isolates of S. Enteritidis tested were assigned to MLST 11, according to the MLST database (http://mlst.ucc.ie/mlst/dbs/Senterica). In previous studies using the same database [12,26,30], MLST was highly correlated to Salmonella serotype. Noda et al [30] found that 30 S. Enteritidis isolates collected in Japan between 1973 and 2004 had homologous MLST type 11 sequences and no nucleotide differences in seven housekeeping genes. Another study targeting seven different housekeeping and virulence genes found that MLST was not able to discriminate clinically relevant serotypes of Salmonella well [38]. The limited discriminatory ability of MLST may be a result of the moderate to slow rate of mutation accumulation within the targeted housekeeping genes [13]. Therefore, the discriminatory performance of MLST needs to be increased if more variable gene targets are examined.
- To the best of our knowledge, this study is the first in which the utility of the five subtyping methods were compared to differentiate S. Enteritidis isolates from both humans and food sources. The present report describes the preliminary results of the use of a small number of S. Enteritidis isolates from sporadic cases. Further research is required on the use of a number of S. Enteritidis isolates that have various epidemical characteristics. A combination of various typing methods may increase the ability to discriminate among similar serotypes of S. Enteritidis and to determine the origin of pathogens. This, in turn, may lead to improved source tracking of foodborne pathogens during outbreaks.
Discussion
-
Acknowledgements
- This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MEST) (No. 2011-0005808). The first author and Jung-Whan Chon were also partially supported by the Brain Korean 21 (BK21) Project from the Ministry of Education and Human Resources Development. We gratefully acknowledge the assistance of Yun-Gyeong Kim, Chang-Hyun Seong, and Dong-Hyeon Kim.
- 1. Zou M, Keelara S, Thakur S. Molecular characterization of Salmonella enterica serotype Enteritidis isolates from humans by antimicrobial resistance, virulence genes, and pulsed-field gel electrophoresis. Foodborne Pathog Dis 2012;3;9(3). 232−8. PMID: 22283616.ArticlePubMed
- 2. Voetsch AC, Van Gilder TJ, Angulo FJ, et al. FoodNet estimate of the burden of illness caused by nontyphoidal Salmonella infections in the United States. Clin Infect Dis 2004;4 15 38(Suppl. 3). S127−34. PMID: 15095181.ArticlePubMed
- 3. Angulo FJ, Tippen S, Sharp DJ, et al. A community waterborne outbreak of salmonellosis and the effectiveness of a boil water order. Am J Public Health 1997;4;87(4). 580−4. PMID: 9146435.ArticlePubMedPMC
- 4. Cianflone NFC. Salmonellosis and the GI tract: more than just peanut butter. Curr Gastroenterol Rep 2008;8;10(4). 424−31. PMID: 18627657.ArticlePubMedPMC
- 5. Schonenbrucher V, Mallinson ET, Bulte M. A comparison of standard cultural methods for the detection of foodborne Salmonella species including three new chromogenic plating media. Int J Food Microbiol 2008;3;123(1–2). 61−6. PMID: 18192050.ArticlePubMed
- 6. Foley SL, Lynne AM, Nayak R. Molecular typing methodologies for microbial source tracking and epidemiological investigations of Gram-negative bacterial foodborne pathogens. Infect Genet Evol 2009;7;9(4). 430−40. PMID: 19460308.ArticlePubMed
- 7. Lukinmaa S, Nakari UM, Eklund M, et al. Application of molecular genetic methods in diagnostics and epidemiology of food-borne bacterial pathogens. Apmis 2004;Nov-Dec;112(11–12). 908−29. PMID: 15638843.ArticlePubMed
- 8. Foley SL, White DG, McDermott PF, et al. Comparison of sub-typing methods for differentiating Salmonella enterica serovar Typhimurium isolates obtained from food animal sources. J Clin Microbiol 2006;10;44(10). 3569−77. PMID: 17021084.ArticlePubMedPMC
- 9. Swaminathan B, Gerner-Smidt P, Ng LK, et al. Building PulseNet International: an interconnected system of laboratory networks to facilitate timely public health recognition and response to food-borne disease outbreaks and emerging foodborne diseases. Foodborne Pathog Dis 2006;Spring;3(1). 36−50. PMID: 16602978.ArticlePubMed
- 10. Weigel RM, Qiao B, Teferedegne B, et al. Comparison of pulsed field gel electrophoresis and repetitive sequence polymerase chain reaction as genotyping methods for detection of genetic diversity and inferring transmission of Salmonella. Vet Microbiol 2004;6;100(3–4). 205−17. PMID: 15145499.ArticlePubMed
- 11. Beranek A, Mikula C, Rabold P, et al. Multiple-locus variable-number tandem repeat analysis for subtyping of Salmonella enterica subsp. enterica serovar Enteritidis. Int J Med Microbiol 2009;1;299(1). 43−51. PMID: 18706857.ArticlePubMed
- 12. Ben-Darif E, De Pinna E, Threlfall EJ, et al. Comparison of a semi-automated rep-PCR system and multilocus sequence typing for differentiation of Salmonella enterica isolates. J Microbiol Methods 2010;4;81(1). 11−6. PMID: 20114063.ArticlePubMed
- 13. Harbottle H, White DG, McDermott PF, et al. Comparison of multilocus sequence typing, pulsed-field gel electrophoresis, and antimicrobial susceptibility typing for characterization of Salmonella enterica serotype Newport isolates. J Clin Microbiol 2006;7;44(7). 2449−57. PMID: 16825363.ArticlePubMedPMC
- 14. Liu F, Barrangou R, Gerner-Smidt P, et al. Novel virulence gene and clustered regularly interspaced short palindromic repeat (CRISPR) multilocus sequence typing scheme for subtyping of the major serovars of Salmonella enterica subsp. enterica. Appl Environ Microbiol 2011;3;77(6). 1946−56. PMID: 21278266.ArticlePubMedPMC
- 15. Olive DM, Bean P. Principles and applications of methods for DNA-based typing of microbial organisms. J Clin Microbiol 1999;6;37(6). 1661−9. PMID: 10325304.ArticlePubMedPMC
- 16. Hickman-Brenner FW, Stubbs AD, Farmer JJ 3rd. Phage typing of Salmonella Enteritidis in the United States. J Clin Microbiol 1991;12;29(12). 2817−23. PMID: 1757554.ArticlePubMedPMC
- 17. Ward LR, de Sa JD, Rowe B. A phage-typing scheme for Salmonella Enteritidis. Epidemiol Infect 1987;10;99(2). 291−4. PMID: 3315705.ArticlePubMedPMC
- 18. Clinical and Laboratory Standards Institute (CLSI) Performance standards for antimicrobial susceptibility testing; 21st informational supplement. M100–21. Wayne, PA;. 2011.
- 19. Gautom RK. Rapid pulsed-field gel electrophoresis protocol for typing of Escherichia coli O157:H7 and other gram-negative organisms in 1 day. J Clin Microbiol 1997;11;35(11). 2977−80. PMID: 9350772.ArticlePubMedPMC
- 20. Matushek MG, Bonten MJ, Hayden MK. Rapid preparation of bacterial DNA for pulsed-field gel electrophoresis. J Clin Microbiol 1996;10;34(10). 2598−600. PMID: 8880529.ArticlePubMedPMC
- 21. Healy M, Huong J, Bittner T, et al. Microbial DNA typing by automated repetitive-sequence-based PCR. J Clin Microbiol 2005;1;43(1). 199−207. PMID: 15634972.ArticlePubMedPMC
- 22. Kidgell C, Reichard U, Wain J, et al. Salmonella Typhi, the causative agent of typhoid fever, is approximately 50,000 years old. Infect Genet Evol 2002;10;2(1). 39−45. PMID: 12797999.ArticlePubMed
- 23. Didelot X, Achtman M, Parkhill J, et al. A bimodal pattern of relatedness between the Salmonella Paratyphi A and Typhi genomes: convergence or divergence by homologous recombination? Genome Res 2007;1;17(1). 61−8. PMID: 17090663.ArticlePubMedPMC
- 24. Falush D, Torpdahl M, Didelot X, et al. Mismatch induced speciation in Salmonella: model and data. Philos Trans R Soc Lond B Biol Sci 2006;11;361(1475). 2045−53. PMID: 17062419.ArticlePubMedPMC
- 25. Perron GG, Bell G, Quessy S. Parallel evolution of multidrug-resistance in Salmonella enterica isolated from swine. FEMS Microbiol Lett 2008;4;281(1). 17−22. PMID: 18279339.ArticlePubMed
- 26. Torpdahl M, Skov MN, Sandvang D, et al. Genotypic characterization of Salmonella by multilocus sequence typing, pulsed-field gel electrophoresis and amplified fragment length polymorphism. J Microbiol Methods 2005;11;63(2). 173−84. PMID: 16226640.ArticlePubMed
- 27. Parsons SK, Bull CM, Gordon DM. Substructure within Salmonella enterica subsp. enterica isolates from Australian wildlife. Appl Environ Microbiol 2011;5;77(9). 3151−3. PMID: 21378038.ArticlePubMedPMC
- 28. Liu WB, Liu B, Zhu XN, et al. Diversity of Salmonella isolates using serotyping and multilocus sequence typing. Food Microbiol 2011;9;28(6). 1182−9. PMID: 21645818.ArticlePubMed
- 29. Hughes LA, Wigley P, Bennett M, et al. Multi-locus sequence typing of Salmonella enterica serovar Typhimurium isolates from wild birds in northern England suggests host-adapted strain. Lett Appl Microbiol 2010;10;51(4). 477−9. PMID: 20809923.ArticlePubMed
- 30. Noda T, Murakami K, Asai T, et al. Multi-locus sequence typing of Salmonella enterica subsp. enterica serovar Enteritidis strains in Japan between 1973 and 2004. Acta Vet Scand 2011;53:38PMID: 21672260.ArticlePubMedPMC
- 31. Grundmann H, Hori S, Tanner G. Determining confidence intervals when measuring genetic diversity and the discriminatory abilities of typing methods for microorganisms. J Clin Microbiol 2001;11;39(11). 4190−2. PMID: 11682558.ArticlePubMedPMC
- 32. Boxrud D, Pederson-Gulrud K, Wotton J, et al. Comparison of multiple-locus variable-number tandem repeat analysis, pulsed-field gel electrophoresis, and phage typing for subtype analysis of Salmonella enterica serotype Enteritidis. J Clin Microbiol 2007;2;45(2). 536−43. PMID: 17151203.ArticlePubMed
- 33. Kilic A, Bedir O, Kocak N, et al. Analysis of an outbreak of Salmonella Enteritidis by repetitive-sequence-based PCR and pulsed-field gel electrophoresis. Intern Med 2010;49(1). 31−6. PMID: 20045998.ArticlePubMed
- 34. Hunter PR, Gaston MA. Numerical index of the discriminatory ability of typing systems: an application of Simpson’s index of diversity. J Clin Microbiol 1988;11;26(11). 2465−6. PMID: 3069867.ArticlePubMedPMC
- 35. Wise MG, Siragusa GR, Plumblee J, et al. Predicting Salmonella enterica serotypes by repetitive sequence-based PCR. J Microbiol Methods 2009;1;76(1). 18−24. PMID: 18835303.ArticlePubMed
- 36. Hyeon JY, Chon JW, Hwang IG, et al. Prevalence, antibiotic resistance, and molecular characterization of Salmonella serovars in retail meat products. J Food Prot 2011;1;74(1). 161−6. PMID: 21219782.ArticlePubMed
- 37. Fernandez J, Fica A, Ebensperger G, et al. Analysis of molecular epidemiology of Chilean Salmonella enterica serotype enteritidis isolates by pulsed-field gel electrophoresis and bacteriophage typing. J Clin Microbiol 2003;4;41(4). 1617−22. PMID: 12682153.ArticlePubMedPMC
- 38. Sukhnanand S, Alcaine S, Warnick LD, et al. DNA sequence-based subtyping and evolutionary analysis of selected Salmonella enterica serotypes. J Clin Microbiol 2005;8;43(8). 3688−98. PMID: 16081897.ArticlePubMedPMC
References
Figure 1Pulsed-field gel electrophoresis (PFGE) patterns of the digested genomic DNA from food and human Salmonella enterica serovar Enteritidis isolates (n = 20). Six antimicrobial susceptibility patterns, three phage types, four PFGE types (A through D), and seven rep-PCR types (1 through 7) were identified among the 20 isolates. AMP = ampicillin, C = chloramphenicol, CN = gentamicin, CTX = cefotaxim, KF = cephalothin, KZ = cefazolin, S = streptomycin, RDNC = reaction does not confirm, TE = tetracycline.
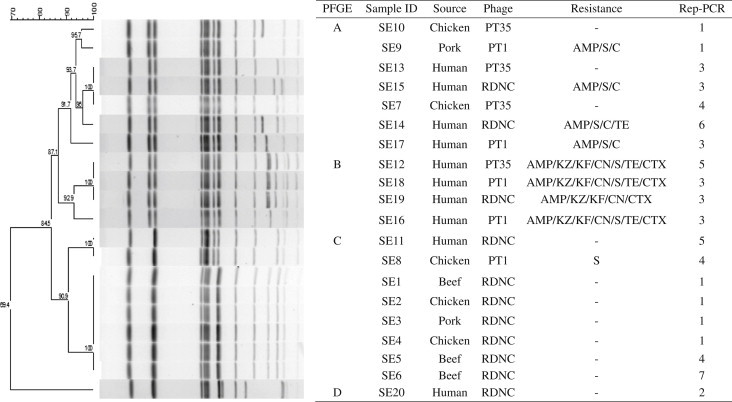
Table 1Comparison of different subtyping methods for Salmonella enterica serovar Enteritidis isolates
| Method | No. of types | Frequent type | Simpson’s index (CI)a |
|---|---|---|---|
| Antibiotic susceptibility | 6 | Non antibiotic resistance | D 0.68 (0.48–0.87) |
| Phage typing | 3 | RDNC | D 0.62 (0.48–0.77) |
| PFGE | 4 | Type C | D 0.71 (0.62–0.79) |
| Rep-PCR | 7 | Types 1 and 3 | D 0.82 (0.73–0.91) |
| MLST | 1 | 11 | D 0 |
Figure & Data
References
Citations
Citations to this article as recorded by 
- Genomic and phylogenetic analysis of Salmonella enterica serovar Enteritidis strains linked to multiple outbreaks in Brazil
Guojie Cao, Maria Balkey, Qing Jin, Eric Brown, Marc Allard, Adma Nadja Ferreira de Melo, Geany Targino de Souza Pedrosa, Tereza Cristina Rocha Moreira de Oliveira, Marciane Magnani, Dumitru Macarisin
Letters in Applied Microbiology.2023;[Epub] CrossRef - Phenotypic and Genotypic Characterization of Salmonella Isolates Recovered from Foods Linked to Human Salmonellosis Outbreaks in Minas Gerais State, Brazil
Leandro L. Faula, Monica M. O.P. Cerqueira, Jovita E. G.C. Madeira, Marcelo R. Souza, Fernando N. Souza, Gláucia C.S. Amancio
Journal of Food Protection.2022; 85(1): 142. CrossRef - Comparison of conventional molecular and whole-genome sequencing methods for subtyping Salmonella enterica serovar Enteritidis strains from Tunisia
Boutheina Ksibi, Sonia Ktari, Houcemeddine Othman, Kais Ghedira, Sonda Maalej, Basma Mnif, Mohamed salah Abbassi, Laetitia Fabre, Faouzia Rhimi, Simon Le Hello, Adnene Hammami
European Journal of Clinical Microbiology & Infect.2021; 40(3): 597. CrossRef - Molecular characterisation of antimicrobial resistance determinants and class 1 integrons of Salmonella enterica subsp. enterica serotype Enteritidis strains from retail food in China
Yu Li, Xiaojuan Yang, Jumei Zhang, Shiyuan Yang, Shuhong Zhang, Moutong Chen, Liang Xue, Yu Ding, Haiyan Zeng, Qihui Gu, Youxiong Zhang, Xianhu Wei, Juan Wang, Qingping Wu
Food Control.2021; 128: 108191. CrossRef - Molecular typing of cephalosporin resistant serovars of Salmonella enterica from poultry and farm animals
A. Rafati Zomorodi, M. Rad, Gh. R. Hashemitabar, H. Salimizand
BULGARIAN JOURNAL OF VETERINARY MEDICINE.2020; 23(2): 178. CrossRef -
Molecular characterization of
Salmonella
isolates recovered from slaughtered poultry in Trinidad
Nitu Kumar, Krishna Mohan, Karla Georges, Francis Dziva, Abiodun A. Adesiyun
Journal of Food Processing and Preservation.2020;[Epub] CrossRef - Effectiveness of BOX-PCR in Differentiating Genetic Relatedness amongSalmonella entericaSerotype 4,[5],12:i:- Isolates from Hospitalized Patients and Minced Pork Samples in Northern Thailand
Kritchai Poonchareon, Chaiwat Pulsrikarn, Narong Nuanmuang, Phichaya Khamai
International Journal of Microbiology.2019; 2019: 1. CrossRef - A common Salmonella Enteritidis sequence type from poultry and human gastroenteritis in Ibagué, Colombia
Luz Clemencia Fandiño, Noel Verjan
Biomédica.2019; 39: 50. CrossRef - Distribution and Molecular Characterization of Campylobacter Species at Different Processing Stages in Two Poultry Processing Plants
Soo-Kyoung Lee, Hyun-Jung Park, Jin-Hee Lee, Jong-Soo Lim, Kun-Ho Seo, Eun-Jeong Heo, Young-Jo Kim, Sung-Hwan Wee, Jin-San Moon
Foodborne Pathogens and Disease.2017; 14(3): 141. CrossRef - Molecular Characterization of Salmonella from Beef Carcasses and Fecal Samples from an Integrated Feedlot and Abattoir in Mexico
Diana Ayala, Kendra Nightingale, Claudia Narvaez-Bravo, Mindy M. Brashears
Journal of Food Protection.2017; 80(12): 1964. CrossRef - Repetitive sequence-based PCR fingerprinting and the relationship of antimicrobial-resistance characteristics and corresponding genes among Salmonella strains from pig production
Teerarat Prasertsee, Nattakarn Khantaprab, Panuwat Yamsakul, Pannita Santiyanont, Nipa Chokesajjawatee, Prapas Patchanee
Asian Pacific Journal of Tropical Disease.2016; 6(5): 390. CrossRef - Prevalence, Seasonal Occurrence, and Antimicrobial Resistance of Salmonella spp. Isolates Recovered from Chicken Carcasses Sampled at Major Poultry Processing Plants of South Korea
Soo-Kyoung Lee, Dasom Choi, Hong-Seok Kim, Dong-Hyeon Kim, Kun-Ho Seo
Foodborne Pathogens and Disease.2016; 13(10): 544. CrossRef - Resistance of Strains Producing Extended-Spectrum β-Lactamases AmongSalmonellafrom Duck Carcasses at Slaughterhouses in Three Major Provinces of South Korea
Soo Kyoung Lee, Dasom Choi, Jung Whan Chon, Kun Ho Seo
Foodborne Pathogens and Disease.2016; 13(3): 135. CrossRef - High Occurrence of Extended-Spectrum β-Lactamase-ProducingSalmonellain Broiler Carcasses from Poultry Slaughterhouses in South Korea
Jung-Whan Chon, Hae-In Jung, Min Kuk, Young-Ji Kim, Kun-Ho Seo, Soo-Ki Kim
Foodborne Pathogens and Disease.2015; 12(3): 190. CrossRef - Comparison of four molecular methods to typeSalmonellaEnteritidis strains
Fábio Campioni, André Pitondo-Silva, Alzira M.M. Bergamini, Juliana P. Falcão
APMIS.2015; 123(5): 422. CrossRef - Modified Method of Multilocus Sequence Typing (MLST) for Serotyping inSalmonellaSpecies
Young-Sun Yun, Su-Jin Chae, Hye-Young Na, Gyung Tae Chung, Cheon-Kwon Yoo, Deog-Yong Lee
Journal of Bacteriology and Virology.2015; 45(4): 314. CrossRef - Molecular Characterization of Salmonella Paratyphi B dT + and Salmonella Heidelberg from Poultry and Retail Chicken Meat in Colombia by Pulsed-Field Gel Electrophoresis
Pilar Donado-Godoy, Barbara A. Byrne, Michael Hume, Maribel León, Enrique Pérez-Gutiérrez, Martha J. Vives Flores, Viviana Clavijo, Ángela Holguin, Juan J. Romero-Zuňiga, Ricardo Castellanos, Mcallister Tafur, Woutrina A. Smith
Journal of Food Protection.2015; 78(4): 802. CrossRef - Prevalence Analysis and Molecular Characterization of Salmonella at Different Processing Steps in Broiler Slaughter Plants in South Korea
Hyun‐Jung Park, Jung‐Whan Chon, Jong‐Soo Lim, Kun‐Ho Seo, Young‐Jo Kim, Eun‐Jeong Heo, Sung‐Hwan Wee, Kidon Sung, Jin‐San Moon
Journal of Food Science.2015;[Epub] CrossRef - Characterization of Escherichia coli–Producing Extended-Spectrum β-Lactamase (ESBL) Isolated from Chicken Slaughterhouses in South Korea
Jong-Soo Lim, Da-Som Choi, Young-Jo Kim, Jung-Whan Chon, Hong-Seok Kim, Hyun-Jung Park, Jin-San Moon, Sung-Hwan Wee, Kun-Ho Seo
Foodborne Pathogens and Disease.2015; 12(9): 741. CrossRef - Comparative Analysis of Subtyping Methods against a Whole-Genome-Sequencing Standard for Salmonella enterica Serotype Enteritidis
Xiangyu Deng, Nikki Shariat, Elizabeth M. Driebe, Chandler C. Roe, Beth Tolar, Eija Trees, Paul Keim, Wei Zhang, Edward G. Dudley, Patricia I. Fields, David M. Engelthaler, N. A. Ledeboer
Journal of Clinical Microbiology.2015; 53(1): 212. CrossRef - Prevalence, characterization, and antimicrobial susceptibility of Salmonella Gallinarum isolated from eggs produced in conventional or organic farms in South Korea
Soo-Kyoung Lee, Jung-Whan Chon, Kwang-Young Song, Ji-Yeon Hyeon, Jin-San Moon, Kun-Ho Seo
Poultry Science.2013; 92(10): 2789. CrossRef
 PubReader
PubReader Cite
Cite
